Теория стадийного развития растений и селекция полевых культур (1936)
Знание закономерностей
индивидуального развития растении для селекционеров и генетиков—дело
существенно необходимое. С этой точки зрения яровизация, а
следовательно, и теория стадийности развития многими своими сторонами
входят непосредственно в селекционно-генетическую науку.
В процессе изучения требований
растительных организмов к условиям внешней среды нам пришлось вплотную
заняться вопросами генетики и селекции. Вести сознательно селекционную
работу методом гибридизации можно только на базе знания закономерностей
развития наследственной основы родительских форм растений.
Оценку наследственной основы
растений, в смысле их пригодности для скрещивания с целью выведения
нового требуемого сорта, генетика до сих пор ещё рекомендует
производить по потомству. Если из скрещивания данных двух сортов в
потомстве через несколько поколений обнаружатся растения с желательной
комбинацией наследственной основы в данных условиях выращивания, то,
следовательно, пара родителей для скрещивания в данном случае подобрана
удачно.
Но ввиду сложности и
чрезвычайной громоздкости выращивания и производства наблюдений над
каждым отдельным растением (выщепенцем) многочисленного потомства
каждой скрещённой пары и ввиду того, что селекционер, работая таким
путем, действует вслепую, наудачу, не представляется возможным брать
одновременно в проработку много (например, хотя бы сотни) скрещённых
пар. При этих условиях процент удачи в выборе пары для скрещивания
настолько мал, что не даёт фактически никакой уверенности громадному
большинству селекционеров в их работе.
Естественно, что при таких методах работы почти невозможно говорить о выведении сортов по строго разработанному плану.
Основная причина
недейственности генетических положений и указаний в вопросе о выборе
родительских форм для скрещивания заключается в том, что генетики
готовые признаки, наблюдаемые у растений, выводят прямо непосредственно
из генотипа, из наследственной основы. Они забывают, что как всё
растение, так и отдельные его органы и признаки есть результат развития
организма, а следовательно, результат многочисленных последовательных
видоизменений и превращений всего содержимого клеток (в том числе и
всего содержимого ядра клеток), Развивающийся
[93]
организм включает
(ассимилирует) соответствующие условия внешней среды. Именно таким
путём и идёт развитие как всего организма, так и отдельных его органов
и признаков.
Любой признак, любое свойство,
наблюдаемое нами у растения, обусловлено развитием наследственной
основы данного растения. Наследственная же основа растения сложилась и
складывается в результате видовой и сортовой истории развития данных
растений.
Благодаря разной истории,
разному развитию получились и получаются различные сорта, то-есть
растения с различной наследственной основой. Поэтому растения каждого
сорта (каждого генотипа) требуют своих условий развития; больше того,
каждая стадия развития одного и того же растения, а также развития
каждого органа и свойства данного растения требует специфических своих
условий. В то же время эти условия требуются растительным организмом не
абсолютно определённые, а относительно. Только потому, что растения
требуют не абсолютно, а относительно определённых условий, они и могут
существовать в ежедневно варьирующих условиях поля. Этим объясняется и
то, что в развитии растений одного и того же сорта, высеянных в разных
условиях, наблюдается нередко довольно сильное варьирование в любом
интересующем нас признаке и свойстве.
Практика нередко ведёт оценку
сортов по малейшим различиям в развитии того или иного свойства
растения. Стоит только растениям одного сорта озимой пшеницы в условиях
данного района быть всего-навсего на 2—3° более морозостойкими, как
этот сорт может быть высеваем на миллионах гектаров, а второй сорт,
уступающий первому в морозостойкости всего лишь на 2—3°, не будет
высеваться вовсе. К примеру озимая пшеница Украинка по зимостойкости
превышает озимую пшеницу Пооператорна не больше чем на 2—3°, и это одно
привело к тому, что Украинка распространялась на юге УССР на миллионах
гектаров, Пооператорна же полностью вытеснена. Можно было бы привести
примеры по целому ряду мельчайших различий в том или ином признаке или
свойстве, благодаря которым один сорт в практике высоко ценится, а
другой бракуется. Из-за небольшого различия в развитии того или иного
свойства нередко получаются большие различия в количестве, а также в
качестве урожая.
Для того чтобы узнать, как
пойдёт развитие и, следовательно, каков будет урожай растений нового
сорта в условиях колхозов данного района, в агронауке пока что
существует только один способ, заключающийся в том, что надо данный
сорт высеять и определить по результатам. Многолетний человеческий опыт
и практика шаг за шагом нащупывают ту среду внешних условий, в которых
высеваемые растения дают наилучший хозяйственный результат. Этим путём
в основном выработаны агроприёмы возделывания различных растений, а
также подобраны и подбираются растения (сорта), наиболее отвечающие
хозяйственным требованиям в принятых условиях агротехники. Мы же
считаем, что перед агронаукой стоит задача разработки такой теории,
которая, будучи тесно увязана с практикой, точнее и полнее могла бы
предвидеть поведение растений в изменённых условиях культуры (климат и
агротехника) с целью создания, во-первых, наилучшей агротехники, а
также с целью предвидения поведения растений в данных условиях культуры
при изменённой путём скрещивания (или иным путём) наследственной
основы, для того чтобы резко ускорить работу по созданию необходимых
нам новых форм и сортов растений.
[94]
Вот почему нам необходима разработка теории развития растений.
Зная довольно полно развитие стадии яровизации, мы теперь легко
можем управлять этой стадией. Любой озимый сорт уже можно путём
агротехники заставить развиваться при весеннем посеве как яровой.
Не производя посева, мы можем теперь заранее указать, в каком
районе данная группа новых для этого района сортов пшеницы, ячменя,
овса, ржи и многих других культур будет яровой или озимой.
Мы уже знаем условия,
требуемые для стадии яровизации вообще, а на этой основе после
лабораторного определения можно узнавать и требования, предъявляемые
каждым сортом в отдельности. Зная условия, требуемые природой данного
сорта для развития стадии яровизации, легко уже путём сопоставления
этих требований с наличными условиями в хозяйствах района знать заранее
поведение в этом смысле растений, если они будут высеяны. Чем лучше мы
будем знать условия для развития той или иной стадии растения, а также
тех или иных признаков, тем лучше сможем предвидеть развитие этих
стадий или признаков в новых условиях выращивания растения.
Особенно это необходимо знать
для целей генетики и селекции при создании новых сортов путём
скрещивания. Ведь все стадии, а также признаки и свойства нового
гибридного растения, полученного из слияния отцовского и материнского
наследственных начал, развиваются в новых условиях, часто отличных от
тех, в которых развиваются рядом же высеянные в один и тот же день обе
родительские формы. В этом случае гибриды и родительские формы
развиваются в одной внешней среде, но используют разные условия этой
среды. Стоит только гибридам пшеницы на 3—5 дней раньше приступить к
выколашиванию, нежели выколашиваются растения родительских форм, как
этим самым развитие семян у гибридных растений будет поставлено в
данных полевых условиях в иные внешние условия, нежели развитие семян у
родительских форм (это довольно рельефно можно было наблюдать в
описанном нами опыте с изменением времени выколашивания при
одновременном посеве различных сортов пшеницы).
Зная условия, требуемые для
развития каждым из родителей, для получения у них тех или иных
признаков или свойств, можно предвидеть ход развития и гибридных
растений в данных конкретных условиях выращивания.
И. В. Мичурин, как никто из
агробиологов постигший развитие растительных организмов, не только мог
предвидеть, из каких двух родителей путём скрещивания можно получить
необходимые ему гибридные растения, но он, зная двойственный характер
требований, предъявляемых для развития тех или иных признаков у
полученных от скрещивания растений (благодаря их гибридному
происхождению), мог путём подстановки необходимых условий, путём
удовлетворения одних требований заставлять развиваться у гибридов
нужные свойства и признаки.
Работа гениального
генетика-селекционера И. В. Мичурина, третировавшегося в царской
России, высоко оценена Советской страной. Мичурин дал нам не только
сотни прекрасных сортов яблонь, груш, слив, вишен, винограда и других
культур, но и ценнейший метод селекционной работы.
Наряду с общебиологическими
основами развития, которые мы должны постигать всё глубже и глубже из
учения дарвинизма, печатные работы Мичурина являются для нас лучшим
руководством в селекционном деле.
[95]
В них всегда можно найти
руководящие указания не только для селекционных работ с плодовыми
деревьями, но и с полевыми сельскохозяйственными культурами.
Используя в нашей практической
селекционно-генетической работе действенные указания как самого
Дарвина, так и его лучших последователей дарвинистов, используя
блестящие положения, обобщённые И. В. Мичуриным на основе его
плодотворнейшей селекционной работы, мы будем иметь возможность
совершенствовать и всё больше и больше конкретизировать наши знания о
развитии растения вообще и в особенности того растения, с которым
данному селекционеру приходится вести практическую работу. С позиций
дарвиновской агробилогической основы нужно читать и обобщать
многочисленный и ценнейший фактический материал, изложенный в научных
книгах и статьях по селекции и генетике многих как наших, так и
заграничных авторов. Все положения, —а они в агробиологической науке
нередки, —не дающие нам действенных указаний, отводящие нас в сторону
от дарвиновского общебиологического учения, должны быть нами отброшены.
Только то, что развивает основное ядро эволюционного учения дарвинизма,
пригодно для построения нашей генетико-селекционой науки.
Фактический материал, довольно
хорошо известный селекционерам, будучи освещён с позиции развития
растения, получает иное, более действенное толкование, принципиально
отличающееся от толкования, принятого сейчас многими генетиками.
Мы исходим из того положения,
что как целое растение, так и отдельные его органы, признаки и свойства
получаются в результате развития наследственной основы. Иными словами,
семенное растение развивается из зародыша семени.
Мы уже указывали, что развитие
растения с необходимостью включает соответствующие условия внешней
среды. Без этого нет развития семенного растения. Это не значит, что мы
исходим из общего положения, что вне времени и пространства ничего не
происходит. Это значит, что условия внешней среды являются неотъемлемой
стороной развития семенного растения.
Необходимо помнить, что
зародыш семени, а также и растительный организм могут вступать во
взаимоотношения с любыми условиями внешней среды, но не при всяком
взаимоотношении (то-есть не с любыми условиями) будет происходить
развитие семенного растения от семени и до созревания новых семян.
Поэтому необходимо условиться, что мы будем говорить только о таких
взаимоотношениях растительного организма с условиями внешней среды,
которые приводят к развитию семенного растения.
Развитие растительного
организма состоит из отдельных, последовательно сменяющихся стадий,
отдельных этапов. Разные стадии развития одного и того же растения
(то-есть разные моменты жизни растения) требуют разных условий внешней
среды. Это не значит, что для прохождения какой-либо стадии развития
растения требуются абсолютно определённые внешние условия. Довольно
хорошо известно, что как всё развитие растительного организма от семени
до семени, так и отдельные этапы, стадии развития могут проходить в
относительно разных условиях внешней среды (в относительно разных
условиях температуры, света, влажности и т. д. ).
Если развивающееся растение
может включать относительно разные условия внешней среды, то,
следовательно, и самое развитие может быть
[96]
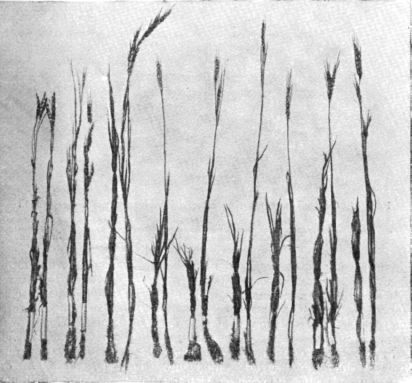
Рис. 42. Часть номеров мировой коллекции пшеницы весеннего посева 1932 г. в совхозе «Гигант», Северный Кавказ.
Каждая пара снопиков—отдельный сорт. Правый снопик каждой пары—посев яровизированным посевным материалом, левый снопик—посев
обычными семенами.
относительно разным. Отсюда, у растений одного и того же генотипа могут развиваться относительно разные качества, свойства и признаки.
Это подтверждают наши опыты по
яровизации мировой коллекции пшениц, ячменя и овса или опыты с посевами
мировой коллекции ячменя в Детском Селе или в Хибинах, о которых
неоднократно в печати и в докладах упоминал академик Н. И. Вавилов.
Путём яровизации, путём сознательного вмешательства лишь в одну из
стадий развития многие сорта пшениц или ячменя, которые до яровизации
не представляли никакой ценности (некоторые из них не давали даже
колосьев), после яровизации оказались намного лучше тех сортов, которые
высеваются в пределах данной области. Генотипы этих растений благодаря
яровизации не изменились, но растения получились разные из
яровизированного и неяровизированного посевного материала. Объяснение
здесь довольно простое. Часть цикла развития, одна из стадий развития
этих растений, именно стадия яровизации, пройдена этими растениями ещё
до посева, в амбаре. Отсюда все остальные этапы (стадии) развития этих
растений хотя и проходили в одном и том же поле с растениями из
неяровизированных семян, но каждая стадия развития растений из
яровизированных семян наступала несколько раньше, чем та же стадия
развития у растений из неяровизированных семян, то-есть развитие
отдельных стадий тех и других проходило в разные дни, хотя и в одном и
том же поле. Этим самым были созданы другие условия для развития этих
растений.
В 1933 г. в Одессе в Институте
селекции и генетики было произведено сортоиспытание 99 сортов ячменя
селекционного отдела. В это сортоиспытание вошли сорта как предыдущей
селекции, так и последних лет. Кроме того, было введено восемь сортов,
взятых с Азербайджанской станции,
[97]
которые без яровизации в
европейской части Союза ССР заведомо непригодны. Наивысший урожай в
этом сортоиспытании получился из «случайно» введённых в сортоиспытание
привозных сортов, высеянных в яровизированном виде.
Этот пример ещё раз говорит,
что в разных условиях может получаться разное развитие растений (разная
конкретизация развития), благодаря чему получаются разные качества и
признаки, вплоть до существенных различий по иммунности. Нам не раз
приходилось наблюдать, как растения, поведение которых (путём
яровизации) изменено, могут па 100% поражаться грибными болезнями, в то
время как контрольные растения, находящиеся с ними в одном и том же
поле, вовсе не поражаются. И, наоборот, не раз можно было наблюдать,
что неяровизированные делянки чрезвычайно сильно поражаются грибными
заболеваниями, в то время как рядом стоящие растения того же сорта из
яровизированных семян вовсе не поражаются. Можно было бы указать ещё
целый ряд других примеров, указывающих на то, что благодаря
относительно разным условиям, которые включаются в разные этапы
развития растения, получаются и различия в органах, признаках и
свойствах растения одного и того же генотипа. Получается разное
количественное выражение признаков,
Рис. 43. Часть номеров мировой коллекции пшениц весеннего посева 1932 г. в Омске.
Каждая пара снопиков—отдельный сорт. Правый снопик каждой пары—из посева яровизированным посевным
материалом, левый снопик—из посева обычными семенами.
[98]
Рис. 44. Озимый ячмень Паллидум 0419. Посев в Гандже весной 1928 г.
органов и даже разное
поведение растения как семенного. Например, один и тот же сорт, при
весеннем посеве в одних районах развивается как озимь (растения не
выколашиваются), в других же районах, тоже при весеннем посеве, этот
сорт развивается как яровой. Этот пример после наших многочисленных
опытов теперь более чем известен. Любой сорт можно сделать озимым или
яровым, высевая его в соответствующих условиях. Правда, для многих
сортов придётся создать искусственную обстановку. Возьмем для примера
такой признак, как мучнистость или стекловидность зерна. Один и тот же
сорт, в один день высеянный, в одном и том же поле может дать
совершенно разную картину в отношении признака мучнистости или
стекловидности зерна в зависимости от времени колошения растений этого
сорта. Каким же образом это получается? Дело в том, что разного времени колошения одного и того же сорта, посеянного в один
[99]
Рис. 45. Просо.
От посева до созревания
выращивалось в условиях непрерывного освещения. Растения двух левых
вазонов из обычных семян к выбрасыванию метёлки не приступили. Растения
двух правых вазонов из яровизированного посевного материала выбросили
метёлку.
и тот же день на одном и том
же поле, мы достигаем тем, что одним растениям мы создаём условия для
быстрого прохождения стадии яровизации (предпосевная яровизация),
другим растениям создаём условия для более быстрого прохождения
световой стадии, третьим—создаём условия для быстрого прохождения как
стадии яровизации, так и световой стадии, четвёртому варианту растений
предоставляем нормально развиваться, то-есть они имеют длину
вегетационного периода «нормальную». Таким путём мы добиваемся того,
что растения при одновременном посеве приходят к разновременному
колошению. Поэтому развитие одних и тех же этапов жизни этих растений
проходит хотя и в одном и том же поле, но в разные дни и,
следовательно, в относительно разных условиях температуры, влажности и
т. д.
В таком опыте, проведённом
нами в 1933 г., растения пшеницы дали урожай зерна различной крупности,
различной окраски семян. Они имели различный внешний вид. В этом опыте
у нас было 3 500 сортов ВИРовской коллекции пшениц. Громадное
большинство этих сортов при ускоренном развитии в районе Одессы в 1933
г. дало хорошо налитое зерно. Вес 1 000 зёрен доходил до 60—70 г. При
обычном же развитии все эти сорта дали значительно худшее зерно, чем
наши стандартные яровые сорта. Стандартные же сорта наших яровых
пшениц, когда мы им помогали в прохождении световой стадии, также
улучшили качество налива зерна. Гирка 0274 (сорт селекции Одесской
селекционной станции) при нормальном развитии дала абсолютный вес 17, 8
г, при ускорении на 6—8 дней развития абсолютный вес зерна получился 28
г.
Как видно из таблицы 1, из 879
сортов пшениц, взятых нами из этого опыта, в результате ускорения
выколашивания всего только 38 сортов не увеличили веса 1 000 зёрен.
Увеличение веса 1 000 зёрен от 1 до 5 г из 879 сортов дали 78 сортов,
от 6 до 10 г—206 сортов, от 11 до 15 г—199 сортов, от 16 до 20 г—216
сортов и от 21 до 25 г—142 сорта.
[100]
Таблица 1
Ускорение выколашивания (в днях)
Количество сортов, ускоривших выколашивание
Количество сортов, не увеличивших веса 1 000 зёрен
Количество сортов, увеличивших вес 1 000 зёрен
от 1 до 5 г
от 6 до 10 г
от 11 до 15 г
от 16 до 20 г
от 21 до 25 г
1
—
—
—
—
—
—
—
2
2
—
1
1
_
_
_
3
13
2
3
4
1
2
1
4
22
2
6
4
7
2
1
5
58
3
10
28
9
4
2
6
98
12
10
36
23
16
3
7
122
8
14
30
31
27
12
8
157
7
15
38
44
39
14
0
150
1
10
33
37
40
29
10
195
1
14
16
21
29
24
11
53
1
4
6
12
20
10
12
39
1
—
8
6
14
10
13
22
—
—
2
2
6
12
14
22
—
—
—
2
10
10
15
26
—
1
—
4
7
14
979
38
88
206
199
216
142
Наконец, несколько слов можно
сказать о признаке остистости. На рисунке 46 представлено на одном и
том же стебле два колоса ячменя. Основной стебель безостый, а боковой
развивался более поздно в иных условиях, чем основной стебель, и
получился остистым.
Приведённых примеров
достаточно для того, чтобы притти к заключению, что любой признак,
орган или свойство растения получаются в результате развития растения.
Развитие же растения проходит через взаимоотношение растительного
организма с условиями внешней среды. Внешние условия включаются
развивающимся организмом как необходимая сторона видоизменений и
превращений, происходящих в растении.
Взаимоотношение же
растительного организма может быть с относительно разными условиями
внешней среды, отсюда самое развитие и его результат могут быть
относительно разными. При относительно разном развитии у растений
одного и того же сорта получаются и относительно разные признаки,
свойства и качества. С тем положением, что растения одного и того же
сорта, развиваясь в разных условиях, дают разные признаки и свойства,
многие из селекционеров и генетиков соглашаются, но с небольшой
оговоркой, что это, мол, касается только тех признаков, о которых
говорилось выше, и может быть ещё некоторых. Но, мол, есть такие
признаки, которые всегда выявляются, которые никогда не изменяются.
Такие признаки, по мнению этих учёных, зависят исключительно от
наследственной основы, и ничего, мол, общего с условиями развития не
имеют. В подтверждение вышесказанного, нередко приводятся примеры того,
когда растения выращиваются в разных географических пунктах, но всё же
эти растения во всех этих пунктах имеют некоторые одинаковые признаки,
другие же признаки, мол, легко меняются. Мне кажется, что эти товарищи
смешивают несравнимые вещи: окружающую растение
[101]
Рис. 46. Растение ячменя. Колос справа— безостистый, колос слева—позже
развившийся, остистый. Ячмень выращивался в Хибинах акад. И. Г. Эйхфельдом.
среду и условия среды,
используемые развивающимся растением. Часто бывает так, что
исследователь довольно сильно изменяет среду, в которой находятся
исследуемые растения, и в то же время вовсе не изменяет тех условий,
которые участвуют в развитии как целого растения, так и того органа или
признака, который исследователь имеет в виду. В таких случаях
исследователь ошибочно приходит к заключению, что развитие данного
признака не связано с условиями внешней среды. Наоборот, бывает и так,
что исследователь как бы не изменяет среду; ему кажется, что она
(среда) почти не изменена, в то же время условия, участвующие в
развитии всего растения или отдельных его органов, в сильной степени
изменены, в результате чего может получиться иное развитие как всего
растения, так и отдельных его органов. Рисунок 27 иллюстрирует отличие
одних растений ячменя от других. Одни растения колосятся, другие же не
имеют стеблей. А ведь это растения одного И того же сорта ячменя,
выращенные на одном и том же поле из обычных неяровизурованных семян,
разница только в том, что одни растения были высеяны в поле 12 марта,
другие тут же рядом высеяны 14 марта. Разница в посеве в два дня
коренным образом изменила поведение растений. Мы теперь уже хорошо
знаем, в чём здесь дело, от чего зависит такое различное поведение
растений в этой, как бы неизменной, среде. Оно зависит от того, что для
одних растений в течение двух дней за весь период вегетации температура
была на 2—3° более низкая.
Следовательно, неправильным
является тот взгляд, что развитие хотя бы и не всех, но некоторых
признаков не зависит от условий среды. Это неверно, так же как
неверно думать, что развитие признаков не зависит от наследственной
основы растения, развивающегося из зародыша семени.
Всё растение со всеми его
признаками и свойствами получается только в результате его развития.
Развитие же семенного растения с необходимостью включает условия
внешней среды.
На базе общего в развитии
семенного растения, которое, как теперь известно, протекает стадийно,
идёт развитие отдельных органов, признаков и свойств растения. Развитие
этих последних тоже идёт при взаимоотношении исходного «внутреннего» с
относительно определёнными внешними условиями. Благодаря относительно
разным внешним условиям (разное питание, увлажнение, освещение и т.
д.), включаемым развивающимися органами, могут получиться и
относительно разные органы и признаки растения (разные количественные и
качественные выражения).
[102]
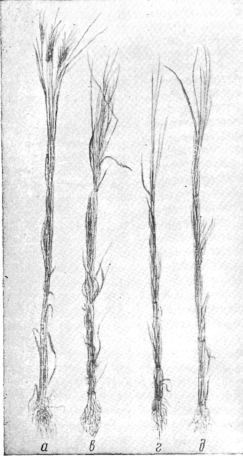
Рис. 47. Растения снопиков а и г.
Эритроспермум 534/1 Азербайджанской станции высеяны яровизированным
материалом (без яровизации растения этого сорта чрезвычайно
позднеспелые). Растения в и д Гирка Одесской
селекционной станции высеяны обычными семенами (предпосевной яровизации
в условиях района Одессы этот сорт не требует).
Левые снопики обоих сортов а и в
выращивались в условиях нормального освещения. Правые снопики обоих
сортов выращивались в условиях беспрерывного освещения на протяжении 25
дней после появления всходов, всё остальное время— в условиях
нормальной продолжительности дня. При прохождении световой стадии в
условиях нормальной длины дня растения снопика о намного обгоняют в
скороспелости Гирку 0274 (снопик в). При прохождении световой
стадии в условиях непрерывного освещения—обратная картина: Гирка (снопик д) обгоняет в развитии
Эритроспермум 534/1 (снопик г). Сорт Эритроспермум 534/1 в условиях района Одессы—позднеспелый,
благодаря стадии яровизации. Сорт Гирка в условиях района Одессы—позднеспелый, благодаря световой стадии. Эти два сорта могут
быть родительской парой для скрещивания с целью выведения раннеспелки. Правые снопики г и д)
сняты с поля на 15 дней раньше левых снопиков а и в. Результат см. на рисунке 48.
Та или иная стадия в общем
пути развития семенного растения является исходной базой, исходным
«внутренним» для развития тех или иных органов и свойств, которые в
свою очередь могут являться исходной базой, «внутренним» для развития
других органов и свойств растения.
Необходимо твёрдо помнить: 1)
каждая стадия развития растения, а также развития каждого органа
требует своих условий внешней среды, но эти специфические свои условия
требуются растениями не абсолютно определённые, а относительно
определённые: 2) разные генотипы, разные сорта для прохождения
однотипных стадий (например, яровизации) или для развития однотипных
органов требуют также относительно разных условии.
Задача селекции, ведущей свою
работу методом гибридизации, состоит в том, чтобы получить такой
генотип, который в процессе индивидуального развития растения дал бы
желательные формы растений именно в условиях колхозов и совхозов
данного района, а не для какого-либо другого района, для которого
селекционер и не думал выводить сорт. До сих же пор часто бывало и так,
что сорт выводился для данного района, а распространение получал в
другом. Например, Гордеиформе 010, Днепропетровской станции, на
территории УССР не имеет ни одного гектара. Этот сорт на значительных
площадях высевается в Сибири и Зауралье. Кооператорка, выведенная
Одесской станцией на территории УССР, имеет чрезвычайно
ограниченное распространение, но нашла себе новую родину на Кавказе.
[103]
В Одесской области введены
Украинка, Мироновской станции, Киевской области, и Гостианум 0237,
Саратовской станции. Наиболее распространённым на территории УССР
сортом яровой пшеницы в настоящее время является Лютесценс 062,
выведенный в Саратове. То же относится к сортам и многих других
культур. Во всех этих случаях селекционером, то-есть лицом, создавшим
данный сорт, больше является не тот, кто создал или отобрал новую
форму, которая оказалась непригодной для тех условий, для которых её
отбирали, а тот, кто из существующего многообразия сортов отобрал
данную форму для культуры в данном районе. Селекционер должен как можно
лучше знать ту среду внешних условий, в которой будут в хозяйствах
развиваться растения селекционируемой им культуры. Он должен знать
климатические условия района и его агротехнические возможности, а также
те требования, которые предъявляются социалистическим хозяйством к
выводимому им сорту. Он должен всё это знать для того, чтобы путём
скрещивания создать такие генотипы растений, требования которых к
условиям внешней среды для развития так совпадали бы с меняющимися
полевыми условиями, чтобы растения в нужные моменты (для данного
района) были в таком состоянии своего развития, в котором они могут
развивать наибольшую стойкость против действия данных неблагоприятных
условий, а также чтобы развитие хозяйственно интересующих нас органов и
признаков сорта происходило, во-первых, в тот период времени, когда
полевые условия наилучше соответствуют требованиям развития этих
органов, и, во вторых, чтобы время созревания урожая наилучше
соответствовало хозяйственным задачам и возможностям.
Таким образом, селекционер,
создавая план-проект будущего сорта, должен по возможности полнее
знать, когда, в какие сроки растения его сорта должны в полевых
условиях проходить разные стадии своего развития и давать урожай. При
этом, как уже указывалось, необходимо, исходя из хозяйственных
требований и заданий, предъявляемых к данной культуре, наилучшим
образом увязать внешние полевые условия района (которые ежедневно в
течение года изменяются) с внутренними требованиями и условиями
развития растения. Увязку необходимо выбрать такую, чтобы растения в
периоды действия неблагоприятных условий были и состоянии наибольшей
выносливости, и в то же время созревание урожая этих растений должно
проходить в условиях, наиболее отвечающих требованиям развития тех
частей и органов растений, ради которых они культивируются. Установив
хотя бы по главнейшим (для данной культуры и данного района) моментам
развития растения календарные даты, в которые растения будущего сорта
при нормальном полевом посеве должны достигнуть определённого состояния
(развития), селекционер и должен теперь путём скрещивания создать такой
сорт, требования которого к условиям внешней среды будут хозяйственно и
биологически наилучше удовлетворены наличными условиями данного поля.
Выше уже приводились примеры,
как у растений одного и того же сорта, то-есть с одной и той же
наследственной основой, при одновременном их посеве в одном и том же
поле может в сильной степени варьировать развитие одних и тех же
свойств, органов и признаков только оттого, что растения одних
вариантов, благодаря предпосевной яровизации или искусственному
удлинению продолжительности дневного освещения в момент
[104]
Рис. 48. Расщепление второго поколения гибридов Эритроспермум 534/1
Азербайджанской станции и Гирка 0274 Одесской станции.
Отдельные выщепенцы из
гибридных растений более раннеспелы, нежели оба родителя:
а—Эритроспермум и б—Гирка. В условиях района Одессы Эритроспермум
задерживается на стадии яровизации, признаком чего является ускорение
созревания после яровизации (растение а1). Гирка задерживается на световой
стадии.
прохождения световой стадии,
всего на несколько дней раньше в условиях поля приступают к развитию
тех или иных органов. Этим мы хотим подчеркнуть хозяйственную и
биологическую значимость продолжительности вегетационного периода
растений в данных полевых условиях.
Агроприём предпосевной
яровизации, дающий возможность управлять временем прохождения одной
только стадии развития растения (яровизацией), для агротехники во
многих случаях имеет большое хозяйственное значение, вплоть до
выращивания в обычных полевых условиях тех растений, которые вовсе не
могли в этих условиях плодоносить, так как это даёт возможность
сокращать вегетационный период растений. Для селекции же способ
создания растений с такой наследственной основой, которая будет иметь
определённую требуемую длину вегетационного периода в условиях данного
поля, имеет несравненно большее значение. Нет культуры и района, где не
требовалась бы та или иная определённая длина вегетационного периода
данного растения. От той или иной длины вегетационного периода растений
зависит, в какие дни, а это значит в
[105]
каких из наличных в районе условий, будут развиваться те или иные органы и признаки растения.
Ввиду того что климатические
условия по отдельным годам довольно сильно в большинстве районов
варьируют, а в связи с этим варьирует и длина вегетационного периода
растений одного и того же сорта, устанавливать точные календарные даты,
в которые растения будущего сорта достигнут того или иного состояния
своего развития, невозможно. Лучше за единицу измерения длины
вегетационного периода намеченного к выведению нового сорта брать не
количество дней вегетации от посева до созревания или, например, у
хлебных злаков от посева до выколашивания, а количество дней, на
которое новый сорт должен запаздывать или обгонять какой-либо из
известных существующих в данном районе сортов.
В нашей работе «Теоретические
основы яровизации» мы старались изложить достигнутые нами небольшие
знания в области общих закономерностей индивидуального развития
семенного растения. Какие же из всего сказанного вытекают практические
выводы?
На основе изложенного о
развитии растений при практическом знании и навыке в селекционном деле
во многих случаях уже можно создавать в короткие планово намеченные
сроки сорта яровых культур с определёнными свойствами и качествами в
условиях данного района.
Основная наша задача
заключалась в выявлении тех условий, которые необходимы различным
генотипам для того, чтобы из высеянных семян получились семена. В
развитии растений нами обнаружена стадийность. Развитие семенного
растения происходит этапами, стадиями. Зная внешние условия, которые
требуются для развития, для прохождения определённой стадии, к примеру
яровизации, оказалось нетрудным делом предсказать, в каких условиях,
то-есть в каких районах любой сорт пшеницы из мировой коллекции при
весеннем посеве будет яровым или озимым. Мы хорошо теперь знаем, что
генотипы (различные сорта, например, пшениц) в озимости и яровости
довольно сильно различаются, хотя признаки озимость и яровость в
генотипе и не заложены; знаем, в каких условиях как поведёт себя любой
сорт в смысле признака озимости или яровости. Этим самым мы изучили
закономерность развития свойства наследственной основы в смысле
озимости и яровости сорта. Наследственная же основа, развиваясь, даёт
не только озимость или яровость. Мы теперь знаем не только стадию
яровизации, но и световую стадию. Известно также, что световая стадия
может проходить (развитие этой стадии может проходить) только после
прохождения стадии яровизации, то-есть исходной базой для световой
стадии являются качественные изменения, свойственные стадии яровизации.
Следовательно, качественные изменения, приобретаемые клетками при
прохождении стадии яровизации, становятся тем внутренним, той основой,
которая при взаимоотношении с относительно определёнными внешними
условиями даёт в дальнейшем те качественные изменения, которые мы
именуем световой стадией.
Дальнейшие стадии в общем
развитии семенного растения, а также развитие отдельных органов
проходят, конечно, таким же путём— путём последовательного
качественного превращения одного содержимого клеток в другое. Не любое
качественное состояние содержимого клеток, а только относительно
определённое может стать непосредственным исходным для превращения в
данное новое качество. Конкретное знание последовательности этих
качественных переходов (что из чего
[106]
развивается), а также знание
условий внешней среди, требуемых отдельными превращениями, и даёт
основание как для предвидения, а отсюда и для управления индивидуальным
развитием данных растений, так и для планового создания путём
скрещивания новых необходимых нам форм растения.
Уже на базе знания двух
выявленных нами стадий развития семенного растения в конце 1932 г. нам
стало ясно, что можно выбирать родительские пары у довольно большого
количества растений (культур), например у пшениц, овса, ячменя,
кукурузы, сои, хлопчатника и целого ряда других культур. Эти выбранные
пары после скрещивания должны давать потомство с определённой длиной
вегетационного периода в условиях данного района, именно в тот
вегетационный период, который селекционеры в селекцируемом растении
хотят получить. Теперь уже в мировой коллекции пшениц, ячменя и овса
можно довольно легко отобрать определённые пары позднеспелых форм,
после скрещивания которых обязательно должны получиться
ультраскороспелые формы растений в условиях данного района (см. рис. 47
и 48).
Можно указать немало пар
среднеспелых форм, например в условиях района Одессы, при скрещивании
которых можно получить позднеспелых выщепенцев. Уже можно лепить путём
скрещивания формы с определённой длиной вегетационного периода для
определённых районов. Для этой цели мировое разнообразие пшеницы,
ячменя и овса, а также и других культур является незаменимым
богатством. На базе мировой коллекции, на базе громаднейшего мирового
разнообразия возможно выбрать пары для создания того или иного
генотипа, который в условиях данного района будет давать определённую
длину вегетационного периода. Для иллюстрации того, как из двух
позднеспелых форм путём скрещивания можно получить раннеспелую,
разберём следующий пример.
Допустим, что в условиях
района Одессы имеются два позднеспелых сорта пшеницы. Один из этих
сортов позднеспелый потому, что он в условиях района Одессы слишком
долго проходит стадию яровизации, все же остальные стадии своего
развития, в том числе и известную нам теперь световую стадию, в
условиях района Одессы этот сорт проходит довольно быстро. Но так как
растения этого сорта задерживаются на стадии яровизации, то общая длина
вегетационного периода будет большая, сорт позднеспелый. Другой сорт л
условиях этого района стадию яровизации проходит быстро, световую же
стадию растения указанного сорта проходят медленно. Все же остальные
стадии своего развития растения этого сорта проходят быстро. В конечном
же результате и этот сорт поздний. Следовательно, оба сорта
позднеспелые, но по разным причинам наследственной основы. Если таких
два сорта скрестить, то в процессе расщепления легко будет отобрать
намного более скороспелые формы, чем оба родителя.
У некоторых генетиков и
селекционеров в январе 1933 г., когда нами впервые был поднят этот
вопрос, явилось сомнение, не будет ли здесь какое-либо «сцепление» или
какая-либо другая премудрость формальной генетики. У многих генетиков
возникло сомнение, можно ли по наследству передать из одного генотипа в
другой те гены, которые в данных условиях дают быстрое прохождение
стадии яровизации, и можно ли эти гены соединять с теми генами, которые
у другого родителя обусловливают в этих же условиях быстрое прохождение
световой стадии. Дальнейшие наши опыты показали, что всё это возможно.
Этим самым
[107]
мы, конечно, ни за, ни против
того, что та или иная стадия определяется двумя или десятью генами.
Любой орган или любой признак определяется всем генотипом. В данном
случае мы этого вопроса не поднимаем потому, что он не является темой
настоящей работы.
Начав в 1933 г. работу по выведению яровой пшеницы на изложенных нами в 1933 г. принципах, мы уже имели к октябрю 1934 г. F6. Работы
по выращиванию родителей начаты в январе 1933 г. Полученные константные
формы от скрещивания двух позднеспелых родительских форм намного более
скороспелы, чем оба родителя, причём не в одной только комбинации, а
почти у всех взятых комбинаций для скрещивания. Всего комбинаций
пшеницы нами взято 30. Есть, правда, в первом гибридном поколении и
небольшие исключения, которые объясняются тем, что длина вегетационного
периода складывается не только из выявленных нами стадий развития.
Например, у некоторых комбинаций скрещивания хлопчатника первое
гибридное поколение даёт слишком сильный рост (жирование). Это
задерживает развитие бутонов. У других растений мощное развитие первого
гибридного поколения в той или иной степени также может отразиться на
задержке цветения и созревания. Ясно, что после выявления ряда других
сторон развития, а не только стадии яровизации и световой стадии и эти
случаи можно будет заранее предвидеть.
Опыты 1933 г. полностью
подтвердили наши предположения о возможности выбирать родительские
формы по стадии яровизации и световой стадии для скрещивания с целью
получения новых форм с определённой, заранее намеченной длиной
вегетационного периода в условиях данного района.
Всё время подчёркиваем: в
условиях данного района, потому что выявленные нами пары могут быть
пригодными (как генотипы для скрещивания) только для условий того
района, в котором их выбирали. Ясно, что в другом районе сами-то
родители могут быть вовсе не позднеспелыми. Мы имеем немало фактов,
когда длина вегетационного периода сильно меняется в зависимости от
района, где эти пшеницы высеяны. Как правило, многие финляндские
пшеницы в сравнении с индийскими в условиях Ганджи выколашиваются на
15—20 дней позже. Эти же самые пшеницы в условиях района Одессы имеют
меньший разрыв, всего на 3—5 дней. В условиях Хибин разрыв между этими
пшеницами также наблюдается, но с обратным знаком. Раннеспелые
индийские в Гандже, в Хибинах делаются более позднеспелыми, чем
финляндские (табл. 2). Поэтому мы всё время подчёркиваем, что в
выбранных парах можно быть уверенным только в условиях данного района.
Выбирать пары для скрещивания в одном районе с тем, чтобы получить
выщепенцев с определённой длиной вегетационного периода в условиях
другого района, пока что мы не готовы. Высказанное положение о выборе
пар для скрещивания пшеницы с целью создания более раннеспелых форм,
чем оба родителя, полностью подтверждалось и для ячменя, овса и кунжута.
На рисунках 28, 31, 32, 34,
35, 36, 39, 40, 48, 49 и 50 можно видеть гибриды, высеянные
одновременно с обоими своими родителями и давшие выколашивание более
быстро, чем оба родителя, как это и предполагалось в момент выбора
родительских форм для скрещивания. Важнейший вопрос, который для
генетики, игнорирующей развитие, не ясен, а именно вопрос
заблаговременного предвидения, из каких двух родителей можно получить
раннеспелые и позднеспелые формы, нами успешно разрешён. Это
объясняется тем, что мы исходим из развития растений, генетика же, нам
кажется, оперирует исключительно с признаками, то-есть только с готовыми результатами развития.
[108]
Рис. 49. Растения ячменя. Гибриды F1 383/6 Азербайджанской (снопик а) ×
032 Одесской селекционной станции (снопик б).
Гибрид F1 более раннеспел, нежели оба родителя.
Рис. 50. Растения ячменя F1 от скрещивания 330/2 Азербайджанской (снопик а) × 046
Одесской станции (снопик б).
Гибрид более раннеспел, нежели оба водителя.
В районах, где хозяйственное
использование некоторых культур упирается в отсутствие необходимых
ранних полностью вызревающих сортов, например в северных районах РСФСР
(спаржевая фасоль, кукуруза, соя и др. ), в районах УССР (хлопчатник,
кунжут) предлагаемым нами способом селекции с привлечением
соответствующего коллекционного материала можно довольно быстро создать
константные формы растений, вызревающие в этих районах лучше, нежели
существующие раннеспелки. В частности, для выведения раннеспелых сортов
фасоли, сои или какого-либо другого растения так называемого «короткого
дня» необходимо взять большую коллекцию сортов, высеять её в обычных
полевых условиях, например Московской или Ленинградской области, и
после всходов через 7—10 дней устроить растениям укороченный день на
протяжении 20—25 дней. Одновременно надо произвести рядом посев
аналогичных сортов, не устраивая им укороченного дня.
Из опытов физиологов по
фотопериодизму мы знаем, что многие сорта фасоли, сои в условиях района
Москвы при искусственно на протяжении 20—25 дней укороченном дне к
осени успевают вызревать. Среди этих вызревших сортов необходимо
наметить тот, который даст наилучший по количеству и качеству урожай.
Это и будет один из родителей, пригодный
[109]
для скрещивания, ибо этому
генотипу нехватает в условиях данного района только фактора темноты
(слишком коротка ночь для прохождения растением этого сорта световой
стадии).
На контрольном участке, на
котором не устраивался укороченный день, необходимо подыскать другого
соответствующего родителя для скрещивания с уже выбранным на опытном
участке. Необходимо на контрольном участке подобрать такого второго
компонента, который по световой стадии в данных условиях нормально
укладывался бы, то-есть такого, который имеет наследственную основу,
развитие которой в условиях данной продолжительности дня идёт примерно
так же быстро, как и в условиях искусственно укороченного дня.
Различие между генотипами как
по стадии яровизации, так и по световой стадии наблюдается довольно
большое. Чтобы выбрать такие растения, надо иметь в своём распоряжении
действительно мировое разнообразие данной культуры. Но на это,
благодаря экспедициям академика Н. И. Вавилова, селекционеры нашего
Советского Союза жаловаться не имеют никакого основания.
Таким же путём можно повести
работу и с хлопчатником. Есть много сортов хлопчатника, которые
позднеспелы по разным причинам. Выбрав пары хлопчатника, у которых
позднеспелость обусловлена разными причинами, можно путём скрещивания
вывести сорт более раннеспелый, чем оба взятых родителя. Из
соответственно подобранных позднеспелых можно комбинировать формы более
скороспелые, чем стандартные скороспелки данного района.
Выведение сорта хорошей яровой
пшеницы для степи УССР (для условий района Одессы)—дело, несомненно,
сложное. Несмотря на двадцатилетнюю с лишним селекционную работу с
яровой пшеницей в Одессе, до сих пор в районах Одесской области нет ни
одного сорта яровой пшеницы одесской селекции.
После того как у нас начал
оформляться способ выбора родительских пар для скрещивания, стал ясным
путь выведения необходимого сорта яровой пшеницы для условий Одесского
района.
На опыте выведения такого сорта с заранее намеченными свойствами ещё в небывало короткий (21/2
года) планом намеченный срок мы в январе 1933 г. и решили проверить и
дальше разработать как метод выбора родительских пар для скрещивания,
так и самую методику планового выведения новых сортов яровых культур.
Опыты, проведённые с посевом
мировой коллекции пшениц в яровизированном и обычном виде, показали,
что для условий района Одессы необходим раннеспелый сорт яровой
пшеницы. Этот сорт должен быть на 5—7 Дней более ранним, нежели
Лютесценс 062 Саратовской селекционной станции, рекомендуемый для
Одесской области.
То, что мы здесь говорим,
конечно, не означает, что всякий раннеспелый сорт будет хорош. Это
только означает, что любой сорт в условиях района Одессы при более
позднем созревании будет хуже, чем при более раннем. Исходя из этого
среди мировой коллекции пшениц по известным нам двум стадиям развития
мы и решили выбрать такие пары, которые при скрещивании дадут
желательные нам раннеспелки. Для скрещивания с целью выведения сорта
необходимо брать не любую пару, которая при скрещивании даст нам только
раннеспелку. Брать нужно только те пары, которые после скрещивания
дадут нам выщепенцев, желательных по длине вегетационного периода и по
всем другим свойствам и признакам.
[110]
Комбинировать же формы для
получения гибридов с определённой длиной вегетационного периода мы были
в состоянии, потому что нами изучены две стадии развития семенного
растения (изучено взаимоотношение развивающегося на этих двух стадиях
организма с условиями внешней среды). После выявления и изучения других
сторон развития растений ещё легче будет подбирать пары для скрещивания
с целью получения выщепенцев, имеющих определённую длину вегетационного
периода d данном районе.
Те пары, из которых можно
скомбинировать путём скрещивания желательный генотип, по длине
вегетационного периода в условиях данного района и которые благодаря
предпосевной яровизации или ускорению прохождения световой стадии
{путём удлинения дня) ещё до скрещивания дают по количеству и качеству
наилучший урожай, и будут компонентными в условиях данного района для
выведения необходимого нам сорта.
Таким образом, на базе
изучения индивидуального развития растения нами выдвинут способ выбора
пар для скрещивания, основанный на выявлении скрытых у родительских
форм возможностей развития на фоне тех полевых условий, для которых
необходимо вывести сорт. Для этого тем или иным агротехническим приёмом
(например, яровизацией или действием на световой стадии) достигают у
растений анализируемых сортов, высеянных в полевых условиях, той длины
вегетационного периода, которая должна быть у будущего сорта в условиях
данного поля. На этом фоне из большой коллекции и выбирают лучшие по
всем хозяйственным показателям пары. Эти пары необходимо комбинировать
так, чтобы требования наследственной основы одного из намеченных
родителей, не удовлетворяемые или плохо удовлетворяемые обычными
полевыми условиями (в чём мы убеждаемся, например, по действию
агроприёма яровизации или по действию удлинённого и укороченного дня в
момент прохождения световой стадии), полностью перекрывались
однотипными требованиями другого намоченного родителя. Требования в
этом направлении другого родителя должны полностью удовлетворяться
обычными полевыми условиями данного района. Узкое же место
наследственной основы другого родителя, которое обнаруживается при
развитии растений в обычных полевых условиях, обязательно должно
перекрываться требованиями наследственной основы первого родителя.
Подбирая таким путём пару для
скрещивания с целью создания нового сорта, можно уже было заранее
предполагать и путь развития в смысле длины вегетационного периода как
растений первого гибридного поколения, так и дальнейших поколений. Эти
предположения на довольно многочисленном опытном материале были
подтверждены, а также получили своё дальнейшее развитие.
Напомним, что для скрещивания
нами выбираются такие компоненты, у одного из которых требования к
условиям внешней среды для развития стадии яровизации плохо пригнаны,
не укладываются или плохо укладываются в наличные условия поля, а
требования для развития световой стадии хорошо укладываются. У второго
же компонента (у второго родителя), наоборот, для прохождения стадии
яровизации данные полевые условия хороши, а для световой стадии плохо
подходят.
Половые клетки (отцовская и
материнская гаметы), образуя зиготу, том самым приносят и все отцовские
и материнские требования к условиям развития.
[111]
Приступая к созданию сорта
яровой пшеницы путём скрещивания выбранных вышеуказанным способом
родительских форм, мы знали, что, начиная со второго поколения, обычно
в результате так называемого расщепления обнаруживается разнообразие
форм в смысле требований их к условиям развития, в результате чего
могут получаться как по своим признакам, так и по поведению разные
растения, в том числе и такие растения, у которых требования к условиям
развития стадии яровизации и световой стадии будут соответствовать
наличным полевым условиям. Неясно было, как же будут в полевых условиях
развиваться растения первого гибридного поколения, зиготы которого
образовались из отцовской и материнской гамет, так как и требования к
условиям развития у этих новых гибридных организмов двойственного
характера, поскольку они разнились в однотипных направлениях у
родительских форм. В частности, требования условий для стадии
яровизации у обоих родителей разнились тоже и для световой стадии.
Прежде всего ещё раз напомним,
что мы понимаем под словами требовать таких-то условий для развития,
например, стадии яровизации. Требовать таких-то условий для развития
стадии яровизации—это значит, что живой белок, содержимое клеток в
таких-то условиях видоизменяется, превращается, приобретает то
качество, которое мы характеризуем стадией яровизации.
Каких же условий для своего
развития потребует растение первого гибридного поколения? Необходимо ли
посевной материал первого гибридного поколения для весенних полевых
условий яровизировать, а также нужно ли для прохождения световой стадии
гибридными растениями искусственное изменение продолжительности
дневного освещения? Ведь растения, представители одной из родительских
форм, в условиях данного поля не могли вовсе яровизироваться или
слишком долго проходили стадию яровизации, с требованием же условий для
световой стадии эти растения легко укладывались в условия данного поля.
Растения другого родителя, наоборот, легко в этих условиях могли
яровизироваться, а с требованием условий для прохождения световой
стадии в обычных полевых условиях данного района, наоборот, плохо
укладывались. В наследственную основу, в зиготу первого гибридного
поколения полностью внесена вся материнская и отцовская наследственная
основа, в том числе и требование условий для прохождения стадии
яровизации, укладывающееся и не укладывающееся в данные полевые
условия, а также и требование условий для прохождения световой стадии,
хорошо укладывающееся (например, от матери) и плохо укладывающееся
(например, от отца) в условия данного поля.
Если искать ответ на эти
вопросы в генетической науке, то мы обычно находим следующее. Потребует
ли в данном случае в полевых условиях первое гибридное поколение
предпосевной яровизации, а также искусственного изменения
продолжительности дневного освещения или не потребует, —будет зависеть
от того, какая из родительских стадий яровизации и световой стадии
доминирует. О том, какая же из родительских стадий в данных полевых
условиях доминирует, генетика даёт такое указание: необходимо высеять
гибриды в поле и посмотреть, какая— отцовская или материнская стадия
яровизации, а также отцовская или материнская световая стадия
превалирует. Та стадия, которая превалирует, та и доминирует. По
существу говоря, никакого ответа на этот счёт генетическая наука не
дала.
[112]
Мы же полагаем, что, зная
развитие той или иной стадии у исходных родительских форм, то-есть зная
условия, которые требуются для прохождения данной стадии каждой
родительской формой, и зная половые условия, в которых будут
выращиваться растения, нетрудно предвидеть и течение развития данных
стадий у гибридного потомства.
Наследственная основа гибрида
развилась из материнско-отцовской наследственной основы путём
биологического слияния двух гамет. Поэтому наследственная основа
гибрида обладает и всеми материнскими и отцовскими требованиями к
условиям внешней среды. Через удовлетворение этих требований из
зародышей семян (или из зиготы) путём последовательных качественных
превращений (то-есть путём развития) содержимого клеток получается
растение со всеми своими органами и признаками.
Следовательно, у гибридных
растений развитие будет проходить на фоне общего последовательного
стадийного развития в направлении лучшего из возможного удовлетворения
требований наследственной основы условиями окружающей растение среды.
Развитие будет проходить по
всем тем возможным направлениям, потребность которых в условиях внешней
среды удовлетворена лучше, нежели удовлетворена этими же условиями
потребность развития растения в однотипном, но всё же в несколько ином
направлении.
Вышесказанное послужило
основой для предположения о том, что развитие первого гибридного
поколения идёт по всем тем возможным для данной наследственной основы
направлениям, требования которых к условиям развития наилучше
удовлетворяются наличной окружающей растение внешней средой.
Поэтому если полевые условия
больше соответствуют прохождению стадии яровизации у растений данного
сорта, нежели у растений другого сорта, то гибридные растения
(полученные от скрещивания растений указанных двух сортов) в этих же
полевых условиях будут так же проходить стадию яровизации, как и
растения первого из указанных родителей.
То же относится и к
прохождению световой стадии. Если один из родителей не может проходить
или плохо, медленно проходит световую стадию в данных условиях, а
другой—хорошо, быстро её проходит, то и первое поколение гибридов будет
проходить световую стадию так же быстро, а в некоторых случаях ещё
быстрее, нежели растения второго родителя. Более быстрое, в сравнении
не только с одним, но с обоими родителями, прохождение световой стадии
у гибридов возможно потому, что данные внешние условия могут
соответствовать для прохождения световой стадии гибридных растений
лучше, чем для растений обеих из родительских форм. Ведь мы знаем, что
в полевых условиях та или иная продолжительность прохождения световой
стадии обычно обусловливается интенсивностью и продолжительностью
дневного освещения, а также температурным фактором. Мы также знаем, что
отношение разных сортов к комплексу внешних условий для прохождения
одной и той же стадии развития (в данном случае световой) разное.
Нередки случаи, когда данные полевые условия для более быстрого
прохождения световой стадии растениями данного сорта пшеницы необходимо
изменить в сторону удлинения дневного освещения, причём эффект здесь
больший, нежели от повышения или понижения температуры. Для растений же
другого сорта, наоборот, требуется изменение температурных условий,
чтобы обеспечить более быстрое прохождение световой стадии, а изменение
продолжительности дневного освещения для растений этого сорта даёт
несравненно меньший эффект.
[113]
Понятно, что первое гибридное
поколение от скрещивания таких родителей по своим требованиям для
прохождения световой стадии будет более пригнано к наличным полевым
условиям и по фактору температуры, и по фактору продолжительности
дневного освещения. В этих случаях гибриды будут проходить световую
стадию при тех же условиях более быстро, чем растения родительских форм.
Наблюдения над растениями
первого поколения гибридов, родительские формы которых подбирались по
разным показателям требуемых условий для прохождения как стадии
яровизации, так и световой, подтвердили вышеизложенные предположения.
Если растения одного из
родителей требуют в данных условиях предпосевной яровизации, а растения
другого родителя этого не требуют, то и гибридные растения предпосевной
яровизации не требуют (см. рис. 31, 40, 49, 50). То же и по световой
стадии: если растения одной из родительских форм кунжута для
прохождения световой стадии требуют удлинения одесской ночи, а растения
другой родительской формы не требуют этого и легко мирятся с данной
продолжительностью дневного освещения, то и гибридные растения не
требуют ночи и, как показали проверочные опыты, хорошо проходят
световую стадию в обычных условиях Одесского района (см. рис. 32, 36).
Все эти положения дают
возможность предвидеть направление развития данных гибридных растений в
данных условиях. Необходимо только знать требования к условиям развития
родительских форм и условия, в которых будет выращиваться гибридное
поколение. Чем полнее мы будем знать биологические требования к
условиям развития наших сельскохозяйственных растений, тем лучше и тем
легче можно будет комбинировать пары для скрещивания, всё с большим и
большим предвидением результатов скрещивания. Знать биологию развития
той или иной стадии растения или отдельных его органов—это значит
знать, из какого звена последовательных качественных изменений
содержимого клеток может развиться данная стадия или данный орган, а
также знать те требования, которые развитие этой стадии или органа
предъявляет к условиям внешней среды.
Исходя из вышеизложенных
теоретических установок, к которым мы пришли в процессе работ по
изучению индивидуального развития растения, а также пользуясь
неоднократными указаниями И. В. Мичурина, который в своих работах
неизменно подчёркивает, что у гибридов развиваются те родительские
свойства, которым больше соответствуют внешние условия развития, нами
были проведены тщательные наблюдения за быстротой развития большого
числа гибридных комбинаций различных растений как в первом, так и во
втором и третьем поколениях, в сравнении с одновременно высеянными
растениями родительских форм. Был также просмотрен в 1934 г. большой
гибридно-селекционный материал по хлопчатнику на Кировабадской
селекционной станции (Азербайджан), затем были просмотрены записи длины
вегетационного периода по большому количеству инцухтированного
материала кукурузы в Днепропетровском зерновом институте, а также
просмотрен литературный материал, касающийся длины вегетационного
периода гибридов. В результате всего изложенного мы пришли к таким
выводам.
1. Растения первого
гибридного поколения в основном приступают к цветению одновременно или
раньше растений ранней родительской формы. Если до скрещивания
произведём стадийный анализ, то можно
[114]
заранее предвидеть, будут
ли растения первого поколения более ранними, нежели ранний родитель. К
моменту цветения растения первого гибридного поколения обычно бывают
или более мощными в сравнении с растениями более мощного родителя или
равными с последним по мощности.
2. Благодаря расщеплению
(обеднению возможностями, приспособления к полевым условиям каждого
конкретного отрезка времени) растения-самоопылители во втором, третьем,
четвёртом и т. д. поколениях обычно не бывают более ранними по цветению
в сравнении с исходной гетерозиго-той. То-есть выщепенцы в
дальнейших гибридных поколениях самоопылителей более ранние по
цветению, нежели растения F1, обычно не появляются. Отсюда у
перекрёстников методом инцухта более ранних по цветению, нежели
исходный гетерозиготный куст, выщепенцев обычно также не бывает.
Данные положения, впервые
изложенные нами совместно с проф. И. И. Презентом в книге «Селекция и
теория стадийного развития растения», вызвали довольно много возражений
со стороны теоретиков генетики и селекции. Мы допускали и теперь
допускаем возможные случаи небольшого, а то и значительного
запаздывания с цветением растений первого поколения по сравнению с
растениями одного из более ранних родителей. Допускаем также случаи и
более раннего цветения отдельных выщепенцев во втором и третьем
поколениях по сравнению с растениями первого1 гибридного поколения.
Эти случаи не только не
противоречат и не умаляют практического значения выставленных нами
положений о развитии гибридных растений в смысле их скороспелости, а,
наоборот, подтверждают эти положения. Случаи запаздывания с цветением
растений первого гибридного поколения обусловлены одним и тем же
явлением, что и обычные случаи наибольшей раннеспелости растений
первого поколения. Это явление, впервые установленное И. В. Мичуриным,
к которому пришли и мы в результате своих работ с полевыми
сельскохозяйственными растениями, заключается в том, что у гибридных
растений всегда развивается та из родительских возможностей, которой
условия внешней среды наиболее благоприятствуют.
Мы уже говорили, что
прохождение каждой стадии развития: яровизации, световой и др.,
гибридных растений идёт так же быстро или даже быстрее в сравнении с
растениями того из родителей, у которого данная стадия проходит
быстрее, нежели у растений другого его компонента (родителя). Развитие
идёт по линии наибольшей (из возможной) приспособленности к наличным на
данном отрезке времени условиям внешней среды. Так же, конечно, идёт
развитие и отдельных органов. Если в данных условиях на данной стадии
своего индивидуального развития растения одного из родителей могут
быстрее, мощнее расти, развивать более мощные стебли по сравнению с
растениями другого родителя на этой же стадии их развития в этих же
условиях, то и растения гибридов первого поколения в этих же условиях
будут не менее, а нередко и более мощными к моменту цветения, нежели
растения более мощной родительской формы. Мы знаем, что время цветения
и созревания однолетних растений в основном, но далеко не целиком,
определяется быстротой прохожде-
1 Здесь, конечно, мы не говорим о тех случаях запаздывания, которые обусловлены варьированием (неоднородностью) внешней среды.
[115]
ниястадий развития. В
практике же хорошо известно, что у растений нередко буйный рост, буйное
развитие стеблей (жирование) являются причиной той или иной задержки
развития органов плодоношения1..
Запаздывание цветения
растений первого поколения в сравнении с растением более ранней
родительской формы всегда будет налицо в тех случаях, когда растения
поздней родительской формы в данных условиях обладают свойством
значительно более мощного развития, нежели растения ранней родительской
формы.
Эти случаи запаздывания
цветения растений первого гибридного» поколения, хотя обычно они в
селекционной практике встречаются редко, также можно предвидеть
заранее, ещё до скрещивания, зная развитие-родителей и сопоставив
требования к условиям внешней среды этих. родителей с условиями полевой
среды, в которых будут произведены посевы. Таким же путём можно также
заранее предвидеть и появление более слабых и более ранних выщепенцев в
дальнейших гибридных поколениях, нежели растения первого поколения.
Теоретически установленные и
практически проверенные и подтверждённые нашей селекционной практикой
изложенные здесь закономерности, вскрытые на основе подхода к
наследственному основанию с позиций развития, дают возможность сделать
более сознательным весь селекционный процесс. Исходя из этих
закономерностей, можно всё время контролировать правильность и на ходу
учитывать и исправлять ошибки селекционного процесса.
Для яровых культур (хлебных
злаков, хлопчатника, кунжута, риса и многих других растений) уже можно
не только с несравненно большей, уверенностью выбирать родительские
пары для скрещивания, из расщепления которого при соответствующей
работе можно вывести заранее намеченный сорт, но, что самое главное,
уже можно знать, из каких: гибридных растений, начиная с растений первого поколения, нельзя вывести и намеченного сорта. Это даёт возможность селекционеру вести по определённым показателям массовую полевую браковку гибридного материала.
Поставив себе целью получение
более раннеспелых форм, чем оба; родителя, можно по растениям уже
первого поколения проверить правильность выбора родительских форм. Если
первое поколение будет более позднеспелым, нежели это намечено для
будущего предполагаемого» сорта, то это является верным признаком того,
что при выборе родительских форм произошла ошибка.
Исходя из того что выщепенцы
второго поколения и последующих: поколений не могут быть более
раннеспелыми, чем само первое поколение, можно по первому поколению
производить браковку в смысле длины вегетационного периода. Для этого в
тех случаях, когда основным лимитом хозяйственного использования данной
полевой культуры
1 В садоводстве
нередко практикуют различное агротехническое вмешательство для
прекращения жирования плодовых деревьев. Все эти агротехнические
способы; в основном сводятся к ослаблению мощного развития стеблей,
благодаря чему ускоряется образование органов плодоношения. Из
способов, применяемых в садоводстве, можно указать на такие, как
накладывание на ствол дерева или на отдельные сучья плодовых поясов для
ослабления корневой системы, а также для повышения количества
пластических веществ в стеблях; уменьшение полива, если применяется
искусственное орошение, уменьшение количества вносимого минерального
удобрения и т. д. Как всем известно, прививки на карликовых подвоях
ускоряют плодоношение и уменьшают мощность развивающегося привоя.
[116]
является отсутствие
раннеспелых сортов, успевающих вызревать к осени, мы советуем
производить скрещивание большого числа пар (сотен комбинаций), не
производя тщательного предварительного стадийного анализа. В этом
случае выгоднее ограничиваться грубой оценкой требований растений к
условиям внешней среды и скрестить побольше комбинаций. По поведению
растений первого поколения необходимо произвести самую строгую браковку
всех тех комбинаций, растения которых оказались более поздними, нежели
намеченный будущий сорт. Необходимо оставить только 1 —2 комбинации
(если они окажутся) такие же или более раннеспелые, нежели сорт,
который селекционер задал себе целью вывести. Если ни одной такой
комбинации не окажется, то необходимо снопа произвести посев большого
числа новых комбинаций и опять просмотреть гибридный материал первого
поколения.
В тех случаях, когда многие
сорта данной культуры свободно успевают вызревать (например, хлебные
злаки в зерновых районах СССР) и требуется только определённая длина
вегетационного периода растений для того, чтобы эти растения не
попадали под действие неблагоприятных климатических условий (засуха,
суховей и т. д. ), нельзя вести отбор гибридных комбинаций в основном
только по длине вегетационного периода первого гибридного поколения.
Ведь готовых (уже существующих в коллекции) форм с требуемой длиной
вегетационного периода можно найти немало, и в то же время они, как
правило, как сорта оказываются непригодными по разнообразным причинам.
Поэтому при селекционной работе с такими культурами по длине
вегетационного периода первого гибридного поколения можно только
определённо сказать, что если первое поколение позже намеченного к
выведению сорта, то такую комбинацию необходимо забраковать. Если же
первое поколение по длине вегетационного периода удовлетворяет, так это
ещё не говорит о том, что данная комбинация пригодна для дальнейшей
работы с целью выведения сорта. В этих случаях, как уже указывалось,
селекционеру для большей уверенности в работе необходима оценка всех
хозяйственных показателей родительских форм в полевых условиях на фоне той длины вегетационного периода, какую должен иметь будущий сорт.
Длина вегетационного периода
первого поколения здесь будет говорить не о правильности выбора
родительских форм, а только о правильной разбивке выбранных форм по
парам в смысле создания у гибридов необходимой длины вегетационного
периода.
Для условий района Одессы мы
пришли к выводу, что любой сорт яровой пшеницы, который будет позже,
чем Лютесценс 062 Саратовской станции, непригоден. (Это, конечно, не
значит, что любой раннеспелый сорт будет здесь пригоден. ) Отсюда,
приступив к выведению ярового сорта для условий района Одессы, мы
заранее поставили себе задание, что будущий сорт ни в коем случае не
должен быть более позднеспелым, чем Лютесценс 062. Он обязательно
должен быть более раннеспелым, хотя бы на 2—3 дня. Изложенные здесь
установки дали нам возможность не только производить тщательный выбор
родительских форм пшеницы для скрещивания, но и производить первую
проверку селекционной работы по первому гибридному поколению, чего
обычно до сих пор селекционеры не применяли. Мы у себя, имея в полевом
посеве несколько комбинаций растений F1, хотя и полученных путём скрещивания родителей, изученных по стадиям развития, обязательно проверяем правильность произведённого
[117]
в подборе пар, так как от этого зависит весь дальнейший успех работы.
Необходимо забраковать, если они окажутся, все те комбинации, которые в F1более позднеспелы или одинаковы по длине вегетационного периода с Лютесценс 062.
Необходимо оставить только то комбинации, которые в F1более раннеспелы, чем Лютесценс 062.
Высеяв F2, которое
уже будет расщепляться как по стадиям, так и по органам и признакам,
развивающимся на базе этих стадий (в том числе и по длине
вегетационного периода), в первую голову ещё в поле необходимо
обязательно забраковывать всех тех выщепенцев, которые более
позднеспелы, чем будущий намеченный к выведению сорт. Исходя опять-таки
из того, что, отобрав, например, выщепенцев одинаковых или более
позднеспелых в условиях нашего района, чем Лютесценс 062, так как в
дальнейших поколениях всё равно они не смогут выщепить (не будет
биологического основания для выделения) формы с нужном нам длиной
вегетационного периода, мы дальнейшую селекционную работу, то-есть
отбор кустов, ведём лишь с оставшимся небольшим процентом выщепенцев. В
работах по селекции яровой пшеницы мы в F2оставляем
только те растения, которые более раннеспелы, чем 062, хотя бы на 2
дня. Например, в наших опытах 1934 г. в комбинации Эритроспермум 534/1 ×
0274 (Гирка местной селекции) из 755 растений в F2раньше
Лютесценс 062 выколашивание дали только 176 растений (растения, давшие
колошение до 6/VI), остальные 579 колосились позже и нами забракованы
ещё во время колошения. В комбинации Эритроспермум 1637/1x062 из 176
растений оставлены только 34. В комбинации Эритроспермум 1637/1×0274
(Гирка) из 1 366 растений в F2оставлены по длине
вегетационного периода только 345, остальные забракованы ещё в поле, и
т. д. Сравнительно небольшой процент растений, оставшихся после
браковки по длине вегетационного периода, подвергается уже обычной
селекционной браковке, которая, конечно, тоже намного уменьшит число
кустов (выбрасываются дегенерированные формы, явно непродуктивные и т.
д. ) в зависимости от того или иного подхода и тех требований, которые
предъявляются к выводимому сорту (табл. 2). То же самое производится в F3и F4и т. д., вплоть до получения константной, готовой нужной формы для условий данного района.
Идя изложенным путём, сразу же
можно в работе натолкнуться на такие преграды: родительские формы
выбраны для скрещивания правильно. На фоне длины вегетационного периода
нужного нам сорта они дают и качественно и количественно хороший
урожай. Правильность подбора комбинации подтверждена первым поколением.
Но в дальнейшем в F2или F3они не
дают ни одного выщепенца или чрезвычайно мало выщепенцев таких, которые
имеет смысл оставить (по лимиту длины вегетационного периода) после
произведения браковки указанным нами способом. Селекционеру становится
ясным, что из числа оставшихся форм нельзя вывести сорта, но в то же
время стадийный анализ говорит, что из этой пары родителей сорт может
получиться. Более чем ясно, что в этих случаях выбрасывать такую
комбинацию нельзя. Это говорит лишь о том, что для посева F2взят слишком малый масштаб и что к этой комбинации к посеву F2необходимо вернуться, то-есть повторить этот посев, взяв несколько больший масштаб F1To же самое будет относиться и ко всем последующим поколениям.
[118]
Таблица 2. Колошение растений F2 (1934 г. )
Дата колошения, Комбинации
25 / V
26
27
28
29
30
31
1 / VI
2
3
4
5
Всего к 5/VI
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
Не выколосилось
Всего растений
Ersp. 534/1 × Гирка 0274..
—
—
—
—
—
—
4
11
3
33
5
120
176
31
172
116
35
9
12
3
71
3
34
3
14
4
12
4
—
2
5
—
1
1
1
—
2
—
44
755
Ersp. 1637/1
× Lutesc. 062...
—
—
—
—
1
—
1
5
6
1
4
16
34
—
9
72
8
3
5
2
9
2
3
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
29
176
Ersp. 1637/1
× Гирка 0274..
—
—
—
—
2
—
27
23
51
35
10
197
345
17
403
104
55
35
28
11
92
18
52
—
2
—
9
—
—
—
4
—
1
—
2
—
2
—
186
1366
Ersp. 2522/1 × Lutesc. 062..
—
—
—
—
—
—
6
25
34
26
12
128
231
—
36
128
34
88
18
6
53
8
3
3
—
—
2
1
—
—
—
—
1
1
1
—
—
—
26
640
Ersp. 2522/1 × Гирка 0274..
—
—
—
—
1
—
4
15
44
12
—
123
199
15
121
37
34
5
17
7
49
3
2
5
—
—
5
—
—
—
—
—
4
—
—
—
—
—
24
529
Ersp. 3060/15 × Lutesc. 062
1
—
—
3
2
—
25
93
81
17
7
202
431
1
27
232
38
50
22
12
87
50
5
22
9
—
2
3
—
2
1
—
5
1
—
—
—
—
57
1 057
Ersp. 3060/15
× Гирка 0274
—
—
—
—
—
—
9
15
11
57
1
97
190
186
224
43
32
35
8
6
57
65
11
11
18
1
7
3
1
1
5
3
4
1
4
2
—
1
28
947
Ferrug. 1316/2
× Lutesc. 062
—
—
—
—
—
—
—
—
1
—
1
—
2
5
10
8
—
2
—
—
7
—
2
—
—
—
1
—
—
—
—
—
—
—
—
—
—
—
2
39
Итого ........
—
—
—
—
—
—
—
—
—
—
—
—
1608
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
—
5509
Таким образом, выставленные
нами положения дают селекционеру в практической работе возможность
работать всё время в десятки раз с меньшим числом растений, чем это
было до сих пор (по крайней мере требовалось генетической теорией
гибридологического анализа), и в то же время получается возможность в
десятки раз расширять масштаб, вовлекая в селекционную проработку
комбинации из всего существующего многообразия культуры, сокращая
намного время выведения сорта и площадь питомников. Ввиду же небольшого
числа новых форм, претендующих на сорт, каждая в отдельности из этих
форм получит от селекционера несравненно больше внимания, чем это было
до сих пор, когда селекционер имел дело в предварительных
сортоиспытаниях с сотнями претендентов на будущий сорт.
Необходимо ещё раз
подчеркнуть, что выбор для скрещивания родительских форм по стадиям
развития еще вовсе не означает, что для скрещивания можно выбирать
любые формы, лишь бы из них можно было создать новую форму, стадийно
(по длине вегетации) наилучше пригнанную к условиям данного района.
Работая таким образом, любой
селекционер будет чрезвычайно легко выводить по заданию раннеспелки, но
это вовсе не означает, что эти раннеспелки будут сортами, пригодными
для данного района, где раннеспелость может и является важнейшим
фактором. Каждому селекционеру в его практической работе в большой
коллекции, проанализированной по стадиям развития, надо брать для
скрещивания, как правило, не просто те формы, которые стадийно наилучше
для этой цели подходят. Те или иные формы стадийно могут и подойти для
условий данного района, но эти же формы с прекрасными своими стадиями
развития могут не выдерживать засухи или иных каких-либо климатических
и почвенных условий данного района или не отвечать требованиям, которые
предъявляет практика данной культуре.
Поэтому, выбирая родительские
формы, надо выявить хозяйственные показатели этих форм через их
стадийный анализ в условиях данного же района. Нами в 1933 г.
произведено скрещивание выбранных по стадиям развития 30 пар форм. Но в
то же время ещё до скрещивания было отме-
[119]
чено, что для практических
целей для выведения сорта в Одесском районе лаборатория производит
скрещивание только двух пар форм: Эритроспермум 534/1 × 0274 Гирка
(сорт местной селекции) и Эритроспермум 534/1 × 062 Лютесценс
(Саратовской станции).
Взятый в качестве исходной
формы для получения сорта яровой пшеницы для хозяйств Одесского района
сорт Эритроспермум 534/1 Азербайджанской станции без яровизации в
условиях нашего района чрезвычайно позднеспелый и малоурожайный. После
яровизации этот сорт в условиях района Одессы становится скороспелым и
в течение трёх лет даёт довольно хороший урожай. Гирка 0274 местной
селекции в условиях района Одессы даёт меньший урожай, чем Лютесценс
062 Саратовской станции, причём эта же Гирка на 5 дней более поздняя,
чем Лютесценс 062. Вообще же и Лютесценс 062 и Гирка в условиях района
Одессы малоурожайны, показателем чего является хотя бы то, что в
хозяйствах Одесского района ни один гектар яровой пшеницы не
высевается. В яровом зерновом клину все площади занимаются ячменём. Но
стадийный анализ этих форм указывает, что Лютесценс 062 и Гирка 0274,
будучи ускоренными (по световой стадии) на 6—8 дней, дают хорошие
показатели по выполненности и качеству зерна, то-есть по главнейшим
показателям урожайности яровых пшениц в наших южных районах. Так как
Гирка является местным сортом, то поэтому можно предполагать, что в её
наследственной основе при условии ликвидации ее «узкого места» по
световой стадии налицо большие возможности развития в условиях
Одесского района. Лютесценс 062 хотя и не является местным сортом
(выведен Саратовской станцией), но выведен из Полтавок путём
индивидуального отбора, кроме того, этот сорт довольно большое число
лет уже культивируется в районах Одесской области. Этим и объясняется,
что мы взяли эти две комбинации в проработку для быстрого получения
необходимого нам сорта. Этими двумя комбинациями на первое время мы
себя несравненно больше страхуем, чем другими комбинациями, хотя бы и
более заманчивыми по своим стадиям развития, но не проверенными или
мало проверенными по возможностям развития хозяйственных показателей в
тех условиях, для которых выводится новый сорт.
[120]
Полученные из этих комбинаций
константные формы оказались более ранними по колошению в полевых
условиях района Одессы на 8—10 дней по сравнению с Гиркой (один из
исходных родителей) и на 5—6 дней более раннеспелыми, нежели 062
Саратовской станции (рекомендованный сорт для Одесской области).
Кроме того, как показало
половое сортоиспытание в 1935 г., эти новые сорта 1163, 1055 и 1160
более урожайны не только в сравнении с малоурожайными родительскими
формами, но и в сравнении с лучшим рекомендованным для Одесской области
сортом 062, а также в сравнении с лучшим селекционным номером,
полученным за прошлые годы отделом зерновых Института селекции и
генетики1.
1 Подробное изложение истории выведения этих новых сортов см. в журн. «Яровизация» № 3, 1935 г., статья Д. А. Долгушина.